人胚腎細(xì)胞293T
一. 細(xì)胞起源
293T細(xì)胞源于人胚胎腎細(xì)胞��,經(jīng)腺病毒5(Ad5)DNA片段轉(zhuǎn)化獲得�����,并穩(wěn)定表達(dá)SV40大T抗原(SV40 large T antigen)���,使其支持含有SV40復(fù)制起點(diǎn)的質(zhì)粒高效復(fù)制[1]�。
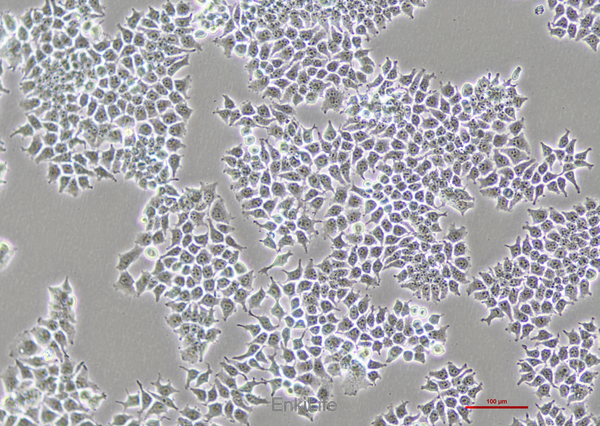
二. 生物學(xué)特性
1. 形態(tài)與生長特性
2. 遺傳與分子特性
-
SV40大T抗原:增強(qiáng)外源基因表達(dá)效率����,促進(jìn)質(zhì)粒復(fù)制[1]。
-
干細(xì)胞樣表型:在無血清三維球體培養(yǎng)中�,高表達(dá)ALDH1、CD44+/CD24-標(biāo)志物��,并上調(diào)β-catenin、Notch1等干細(xì)胞信號(hào)通路[3]����。
-
間充質(zhì)轉(zhuǎn)化傾向:三維培養(yǎng)時(shí)表達(dá)波形蛋白(vimentin)、Zeb1等間充質(zhì)基因�����,可能增強(qiáng)轉(zhuǎn)移潛能[3]����。
3. 代謝與毒性響應(yīng)
三. 培養(yǎng)與儲(chǔ)存
1. 培養(yǎng)基:DMEM+10% FBS+1% P/S[2]。
2. 傳代與凍存:常規(guī)胰酶消化傳代����;30%基礎(chǔ)培養(yǎng)基+60%FBS+10%DMSO,液氮保存[2]�。
3. 轉(zhuǎn)染兼容性:脂質(zhì)體轉(zhuǎn)染效率高(如GFP載體轉(zhuǎn)染成功率50–60%)[7][8]�。
四. 研究應(yīng)用領(lǐng)域
1. 基因功能與調(diào)控研究
-
RNA干擾(RNAi):廣泛用于基因沉默(如p21WAF1/CIP1�����、PES1�、HPIP基因),驗(yàn)證靶基因功能[9][10][11]����。
-
啟動(dòng)子活性分析:如尼羅羅非魚β-actin啟動(dòng)子驅(qū)動(dòng)EGFP表達(dá)[8]。
-
蛋白互作研究:通過免疫共沉淀篩選互作蛋白(如C14orf166與RS8/EFCB/NRAP)[12]����。
2. 疾病機(jī)制模型
-
腫瘤研究:模擬癌干細(xì)胞表型,用于放療抗性�����、轉(zhuǎn)移機(jī)制探索[3]����。
-
代謝疾病:研究AMPK-脂聯(lián)素通路(女貞苷激活A(yù)MPK)[4]�。
-
病毒感染:構(gòu)建禽流感HA-GFP融合蛋白,分析信號(hào)肽對(duì)表達(dá)的影響[7]�����。
3. 藥物篩選與載體開發(fā)
-
抗腫瘤藥物:評(píng)估化合物選擇性(如共軛亞油酸抑制EC109癌細(xì)胞)[5]。
-
基因載體:殼聚糖納米粒轉(zhuǎn)染pGFP���,但存在細(xì)胞毒性[6]�。
-
慢病毒包裝:HPIP siRNA慢病毒抑制HeLa/HepG2增殖[10]���。
五. 近年研究進(jìn)展
1. 自噬調(diào)控:弓形蟲ROP17蛋白在293T中表達(dá)�,促進(jìn)血清饑餓誘導(dǎo)的自噬(LC3-II↑���、P62↓)[13]。
2. 神經(jīng)退行性疾?���。?/strong>帕金森研究中用于驗(yàn)證Parkin蛋白調(diào)控小膠質(zhì)細(xì)胞NLRP3炎癥體[14]。
3. 新型基因工具:構(gòu)建雙標(biāo)簽載體(如pcDNA3.1-Flag-His)用于蛋白互作組學(xué)研究[12]��。
六. 局限性與克服方法
1. 局限性
-
遺傳不穩(wěn)定性:長期傳代可能累積突變���,影響實(shí)驗(yàn)結(jié)果可重復(fù)性����。
-
非完全正常細(xì)胞:表達(dá)SV40抗原及間充質(zhì)基因,不完全代表原代腎細(xì)胞[3]�。
-
轉(zhuǎn)染毒性:殼聚糖納米粒等載體導(dǎo)致細(xì)胞損傷(抑制率>26%)[6]。
2. 克服方法
-
低代次使用:限制傳代次數(shù)(<50代)��,定期鑒定細(xì)胞身份�����。
-
替代載體:優(yōu)化脂質(zhì)體或病毒載體(如慢病毒)降低毒性[10]�����。
-
三維培養(yǎng)模型:利用球體培養(yǎng)模擬體內(nèi)微環(huán)境����,提升生理相關(guān)性[3]。
七. 總結(jié)與展望
293T細(xì)胞因其高轉(zhuǎn)染效率�����、快速增殖及SV40大T抗原特性��,已成為分子生物學(xué)和疾病研究的核心工具�����。近年研究拓展至自噬、神經(jīng)退行性疾病及新型基因編輯領(lǐng)域��。未來方向包括:
1. 精準(zhǔn)基因編輯:結(jié)合CRISPR-Cas9構(gòu)建更復(fù)雜疾病模型����。
2. 類器官整合:聯(lián)合3D培養(yǎng)技術(shù)模擬器官微環(huán)境。
3. 毒性機(jī)制優(yōu)化:開發(fā)低毒載體(如肽納米粒)提升轉(zhuǎn)染安全性���。

參考文獻(xiàn)
1. Viebahn, S. Die Rolle der Oberflächensialylierung bei der Modulation von Immunantworten. 2008.
2. 人胚腎細(xì)胞293T 說明書.武漢恩璣生命科技有限公司.
3. Characterizing cancer cells with cancer stem cell-like features in 293T cells. Debeb BG, et al. Cancer Res. 2010. [PMID: 20663901]
4. 郭文文, 等. 女貞苷通過激活A(yù)MPK促進(jìn)脂聯(lián)素的組裝. 2018.
5. 吳演, 等. 海蓬子籽油亞油酸共軛化產(chǎn)物的抗腫瘤作用. 2012.
6. 王紅梅, 等. 殼聚糖納米?;蜉d體的制備和體外轉(zhuǎn)染實(shí)驗(yàn)研究. 2010.
7. 蘇艷, 等. 信號(hào)肽序列對(duì)禽流感病毒H5N1 HA蛋白GFP融合分子表達(dá)的影響. 20010.
8. 鄒芝英, 等. 尼羅羅非魚β-actin基因啟動(dòng)子的分離及其在真核細(xì)胞中的活性驗(yàn)證. 2011.
9. 王朝云,等.利用RNA干擾抑制p21WAF1/CIP1基因的表達(dá)及周期阻滯效應(yīng)[J].生物技術(shù)通訊,2008.
10. 徐小潔, 等. 慢病毒介導(dǎo)RNA干擾HPIP蛋白抑制腫瘤細(xì)胞增殖. 2011.
11. 李杰萍, 等.利用不同方法檢測RNAi抑制人pescadillo基因的表達(dá)[J].細(xì)胞與分子免疫學(xué)雜志,2007.
12. 張登祿, 等.胰腺癌轉(zhuǎn)移相關(guān)基因C14orf166的真核表達(dá)及其蛋白相互作用的蛋白質(zhì)組學(xué)篩選[J].中國醫(yī)藥生物技術(shù),2010.
13. 劉宏延, 等. 弓形蟲ROP17真核表達(dá)載體的構(gòu)建及ROP17參與自噬的研究. 2012.
14. Parkin regulates microglial NLRP3 and represses neurodegeneration in Parkinson's disease. Yan YQ, et al. Research. 2025.